
Genetik: Hasentiere
Ordnung Hasentiere
Kleine bis mittelgroße Säuger, die in der Ernährungsweise und im Aufbau des Gebisses den Nagetieren ähneln. Ihr Schwanz ist kurz oder fehlt fast völlig. Die sohlen sind behaart. Der knöcherne Gaumen ist stets kurz, dafür sind die Foramina incisiva und die Choanen groß.
Alle Lagomorphen besitzen 2/1 Schneidezähne, die zeitlebens nachwachsen. Die oberen 2. Schneidezähne sind klein und als Stiftzähne ausgebildet (siehe Abb. 2), zwischen Schneide- und Backenzähnen ein großes Diastema. Backenzähne wachsen bei rezenten Arten zeitlebens nach. I1 vorn stets mit Längsrinne. Der Schmelz umgibt die Schneidezähne vollständig.
2 Ordnung Lagomorpha – Hasentiere
-Zur Taxonomie. Die Lagomorpha wurden ursprünglich als Unterordnung Duplicidentata zu den Rodentia gestellt. GlDLEY (1912) hat als erster ihre Eigenständigkeit hervorgehoben und sich dafür ausgesprochen, die Hasenartigen als eigene Ordnung abzutrennen. Dem sind die Taxonomen nach einigem Zögern gefolgt. Sie haben die nächsten Beziehungen bei verschiedenen anderen Ordnungen vermutet, ohne sich entscheiden zu können. LUCKETT (1985) und CHUAN-KUEI et al. (1987) versuchen zu begründen, daß unter den rezenten Säugetieren die Rodentia die nächsten Verwandten der Lagomorpha sind. Hierfür sprechen vor allem Homologien im Schneidezahngebiß (s. S. 8) und Übereinstimmungen im Bau der Plazenta. WADDELL et al. (1999) sprechen sich auf Grund der ihnen vorliegenden Studien mit einer “mittleren Sicherheit” von 50- 80% für ein Schwestergruppen-Verhältnis zwischen Rodentia und Lagomorpha aus; sie lassen die Bezeichnung Glires für das die beiden Gruppen zusammenfassende Taxon wieder aufleben. Schwer zu beurteilen sind zwei fossile, seit dem Palaeozän bekannte Familien, die Eurymyliden und die Mimotoniden. CHUAN-KUEI et al. (1987) zeichnen ein Beziehungsschema, nach dem die Mimotoniden die Schwestergruppe von Lagomorpha und Rodentia wären, die Eurymyliden aber Schwestergruppe der Rodentia allein.
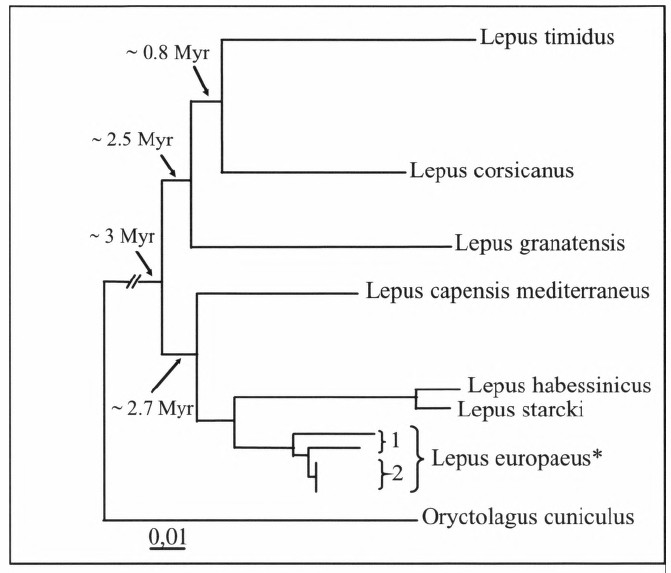
Umfang der Ordnung. Rezent 2 Familien, die Ochotonidae und die Leporidae. Zu den Ochotonidae gehört allein die Gattung Ochotona mit 19 Arten, zu den Leporiden zählen 11 Gattungen mit 46 Arten. Die Artenanzahl ist allerdings in beiden Familien noch etwas umstritten. In Europa sind Ochotoniden zwas bis ins Holozän vorgekommen, heute aber ausgestorben.
Fortpflanzungsperiode: Beginnt in Europa in der Regel im Dezember mit dem ersten Auftreten geschlechtlich aktiver Männchen, bzw. im Januar mit dem Auftreten der ersten trächtigen Weibchen. Endet im September, seltener im Oktober mit den letzten Trächtigkeiten. Nur vereinzelt geschlechtlich aktive Tiere beiderlei Geschlechts bis in den Dezember hinein. Fortpflanzungszyklus photoperiodisch gesteuert (MARTINET 1976). Unter den Bedingungen der Nordhemisphäre resultiert herbstliche Depressionsphase (keine Fortpflanzung, geringstes Gonadenvolumen und -gewicht). Andere exogene (z.B. Witterung, Klima) und endogene Faktoren (z.B. Ernährungszustand) beeinflussen Fortpflanzungsaktivität, können sie verzögern, unterbrechen oder beenden.
Regionale und jährliche Abweichungen von den oben genannten Normwerten für Beginn und Ende der Periode. Bei mildem Klima erste Geburten im Januar, sonst gewöhnlich im Februar, bei kalter Witterung auch erst im März.
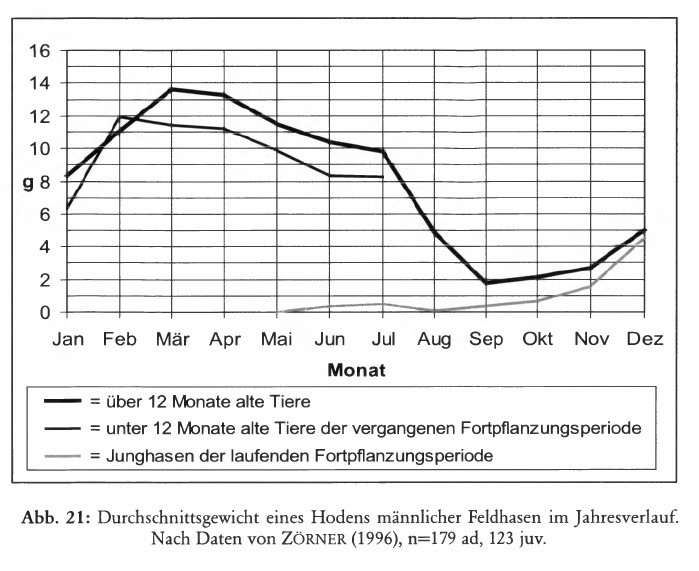
Bei männlichen Feldhasen sind die Gonaden ab September, spätestens in den Monaten Oktober und November in völliger Ruhe. Das Hodengewicht erreicht ein Minimum. Die Hoden sind in diesem Zustand in die Leistengegend zurückgezogen. Die Spermatogenese ist dann unterbunden. Die Hoden sind etwa ab Mitte Dezember wieder in das Skrotum zurückverlagert und sichtbar.
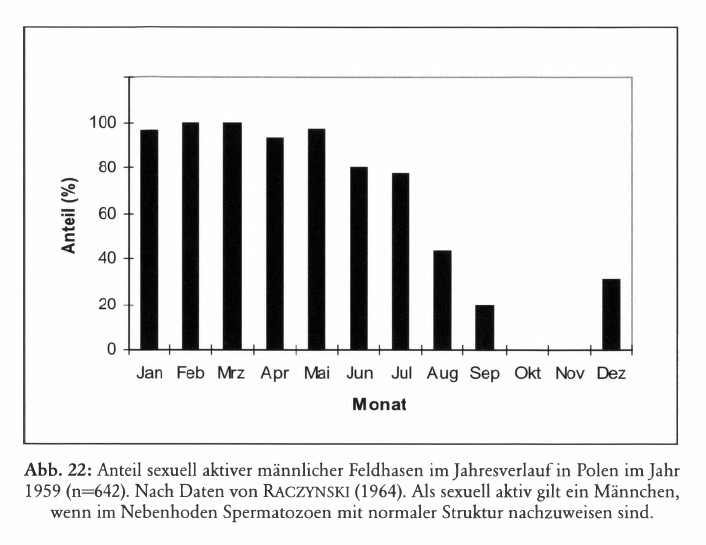
Das durchschnittliche Hodengewicht nimmt ab Dezember rasch zu, es erreicht im März ein Maximum und fällt bereits ab Juli wieder ab (RACZYNSKI 1964, ZÖRNER 1996, Abb. 5). Parallel zur Entwicklung des Hodengewichts ändert sich der Anteil geschlechtlich aktiver Männchen im Jahresverlauf (Abb. 6). ZÖRNER (1996) geht davon aus, dass erst ab einem Testesgewicht von 5 g die Spermatogenese in Gang kommt.
Bei weiblichen Feldhasen ist die Gonadenentwicklung im Jahreszyklus ähnlich, jedoch um einen Monat verschoben. Die Ovarien weisen im November, Dezember das geringste Gewicht und Volumen auf. Erst ab Januar nimmt das Ovargewicht rasch zu und erreicht ein Maximum zwischen Februar und April (RACZYNSKI 1964, ZÖRNER 1996).
Die geschlechtliche Aktivität beginnt und endet somit bei Häsinnen im Durchschnitt etwa einen Monat später als bei den Rammlern. RACZYNSKI (1964) fand in Polen im Jahr 1959 die ersten trächtigen Weibchen im Januar; in der zweiten Januarhälfte waren 20 % der untersuchten Tiere trächtig, im Februar 85 %; die letzten trächtigen Häsinnen wurden im September festgestellt; im Folgejahr traten trächtige Weibchen erst im Februar auf. BROEKHUIZEN und MAASKAMP (1981) fanden in den Niederlanden im Zeitraum 1968 bis 1973 trächtige Häsinnen ebenfalls von Januar bis September. HEWSON und TAYLOR (1975) stellten in Schottland im Rahmen einer 13jährigen Untersuchung folgende Schwankungsbreiten fest: Erste Trächtigkeit 19. Januar bis 28. Februar (Durchschnitt 4. Februar), letzte Trächtigkeit 1. August bis 30. Dezember (Durchschnitt 19. Oktober); die Länge der Fortpflanzungsperiode betrug im Schnitt 259 Tage (bei einer Spannweite von 176 – 344 Tagen). FRYLESTAM (1979) gibt für Schweden an, dass die ersten Geburten im Februar Vorkommen und dass die Fortpflanzung im September, seltener im Oktober endet. HANSEN (1992) fand bei Untersuchungen in Dänemark von 1984 bis 1987 trächtige Häsinnen von Anfang Februar bis Ende August.
Er schätzte nach dem Entwicklungszustand der Embryonen, dass die ersten Geburten Mitte Februar und die letzten Ende September stattfanden. PEGEL (1986) untersuchte die Jagdstrecke des Jagdjahres 1984/85 in verschiedenen Gebieten Westdeutschlands und rechnete vom Augenlinsengewicht der Junghasen auf den Geburtsmonat zurück. Danach stammten auf der Nordseeinsel Föhr die ersten Junghasen aus dem Geburtsmonat Januar, die letzten wurden im September geboren. Auf dem Festland reichte die Spanne von Februar bis Oktober. ZÖRNER (1996) fand in Sachsen-Anhalt (BRD) im Zeitraum 1967 bis 1969 die ersten geschlechtlich aktiven Häsinnen im Januar (5%), im Februar waren es bereits 79 %; die letzten Geburten fielen in den September. MÖLLER (1976) ermittelte für ostdeutsche Bezirke im Zeitraum 1966 bis 1971 eine Länge der Reproduktionszeit von durchschnittlich 236 Tagen (Variationsbreite 215 bis 256 Tage). ZÖRNER (1996) gibt nach eigenen Befunden an, dass die Reproduktionszeit in Ostdeutschland im Durchschnitt 220 Tage andauert und sich jährlich um 1 bis 1,5 Monate verlängern oder verkürzen kann.
Grundsätzlich verläuft der Fortpflanzungzyklus in anderen Erdteilen, wo der Feldhase eingebürgert wurde, nach gleichem Muster, wenn die Verschiebung der Jahreszeiten berücksichtigt wird. Es ergeben sich lediglich mehr oder weniger lange Fortpflanzungsperioden in Abhängigkeit vom Klima (vgl. z.B. BONINO und Montenegro 1997, Dietrich 1985, Flux 1967).
T r a g z e i t: 42 bis 43 Tage. Verkürzung des Abstandes zwischen zwei Geburten durch Superfötation möglich.

Die Tragzeiten können sich zwar unter günstigen Bedingungen nahezu lückenlos aneinanderreihen (RlECK 1956). Jedoch ist das keineswegs die Regel, meist sind Pausen eingeschaltet. Dies folgt allein aus der durchschnittlichen Anzahl der Sätze pro Jahr und der Länge der Fortpflanzungsperiode. Bei Gefangenschaftshaltung ist es aber nicht selten, dass der Abstand zwischen zwei Geburten sogar weniger als 42 Tage beträgt.
Diese Verkürzung wird durch eine Superfoetation ermöglicht. Noch während der Trächtigkeit wird dabei eine erneute Ovulation durch Kopulation ausgelöst. Nach CAILLOL und MARTINET (1983) fuhren Sexualkontakte frühestens ab dem 34. Tag der Trächtigkeit, meist jedoch erst zwischen dem 38. und 40. Tag zu einer „Schachtelträchtigkeit“.
Der Abstand zwischen zwei Geburten wäre demnach nur um wenige Tage geringer als 42. SLAMECKA und SEBOVA (1991) zeigen jedoch, daß in Einzelfällen auch wesentlich kürzere Intervalle zwischen zwei Geburten möglich sind (24 und 27 Tage). Superfoetation kann auch Vorkommen, wenn beide Uterushörner Embryonen enthalten. Die Befruchtung der Eier erfolgt durch überlebende Spermien des vorletzten Geschlechtspartners.
Es ist also nicht erforderlich, dass Spermien aus der letzten Paarung die noch vorhandenen Embryonen passieren müssen (ausführliche Diskussion zum Thema Superfötation siehe u.a. ZÖRNER 1996). Bei in der freien Wildbahn lebenden Hasen kommt die Superfötation aber in der Regel nicht vor (vgl. Raczynski 1964, Broekhuizen und Maaskamp 1981).
Anzahl der Würfe im Jahr: In Europa 1 bis 4, am häufigsten 3 Würfe pro Häsin. Allgemein ist es schwierig, im Freiland die Anzahl der Würfe zu bestimmen, weil die Fortpflanzungsaktivität einem individuellen Muster folgt und nicht synchronisiert ist. Nach FRYLESTAM (1979) schwankt in Schweden die durchschnittliche Anzahl der Würfe von Gebiet zu Gebiet und von Jahr zu Jahr zwischen 2,1 und 3,1. Einen Wurf im Jahr hatten 3 % aller untersuchten Häsinnen, zwei Würfe 8 %, drei Würfe 69 °/o und vier Würfe 14 %. Für Westpolen geht PlELOWSKI (1976a) im Zeitraum 1966 bis 1973 von durchschnittlich 3,2 Würfen pro Jahr aus.
Dies entspricht weitgehend den Ergebnissen von RACZYNSKI (1964), der für das Untersuchungsgebiet in der gleichen Region Polens durchschnittlich 3,4 Würfe pro Häsin annimmt und auf Grund des Anteils trächtiger Häsinnen im Jahresverlauf schätzt, dass der erste Wurf am häufigsten in die Monate März/ April, der zweite im Mai/Juni und der dritte im Juli/August anfällt und dass einige Tiere zum vierten Mal am Ende der Reproduktionsphase werfen. HANSEN (1992) fand in Dänemark im dreijährigen Untersuchungszeitraum 1984 bis 1986 vier Würfe nur in zwei Jahren. In jedem Jahr gab es auch Häsinnen ohne Anzeichen von Fortpflanzungsaktivität (14 bis 21%). Unter Berücksichtigung der unproduktiven Weibchen berechnete HANSEN die durchschnittliche Anzahl der Würfe mit 1,93 bis 2,54 pro Jahr. ZÖRNER (1996) ermittelte für Ostdeutschland in den Jahren 1967 bis 1969 die Durchschnittswerte 3,5; 3,7; 2,4. Für erstmals an der Reproduktion beteiligte Häsinnen nimmt er geringere Durchschnittswerte als für ältere Tiere an. BENSINGER et al. (2000) fanden in westdeutschen Gebieten in den Jahren 1997 und 1998 im Schnitt 4,2 Sätze pro Jahr, bei einer allerdings geringen Stichprobe von 28 untersuchungsfähigen Uteri. ZÖRNER (1996) wertet Literatur aus verschiedenen Ländern aus und zeigt damit regionale, u.a. klimatisch bedingte Unterschiede auf:
Im Süden des europäischen Teils der ehemaligen UdSSR werden drei bis vier Sätze pro Jahr für möglich gehalten, im zentralen Teil jedoch nur zwei; in Rumänien wird mit vier Würfe pro Jahr gerechnet; für Neuseeland, bei längerer Fortpflanzungsperiode werden fünf Würfe pro Jahr genannt. MARTINET (1976) gibt für Käfighaltung durchschnittlich 4,1 Sätze pro Jahr bei natürlichem Lichtzyklus an und 6,9 Sätze pro Jahr unter künstlicher Beleuchtung im Langtag (16L/8D). SLAMECKA und SEBOVA (1991) fanden maximal 6 Würfe pro Jahr bei Käfighaltung, wobei in diesen Fällen die Würfe innerhalb einer Zeitspanne von 183 bis 189 Tagen erschienen und die Intervalle zwischen den Geburten durch Superfötation verkürzt waren (durchschnittliches Intervall 36 bis 38 Tage).
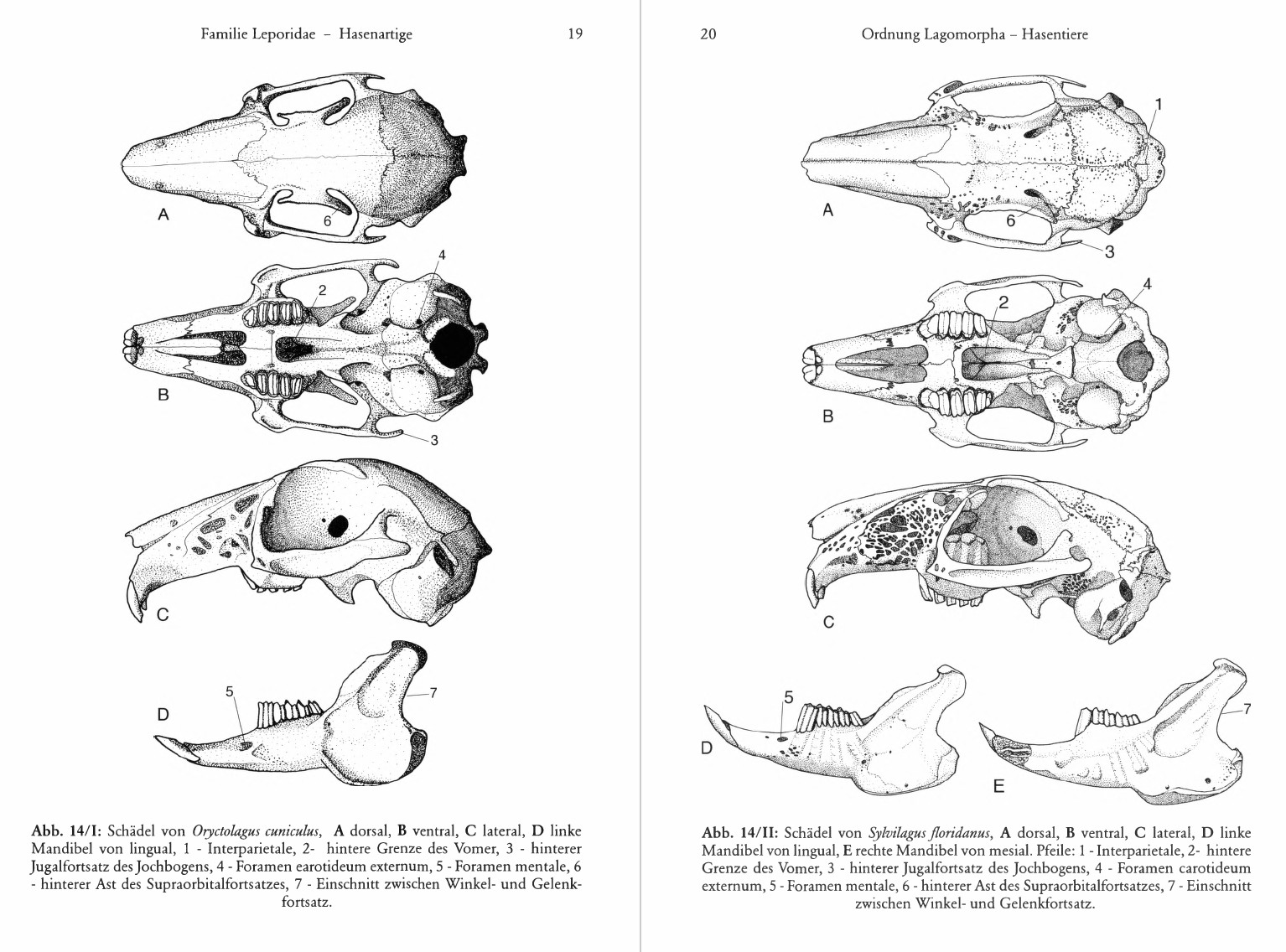
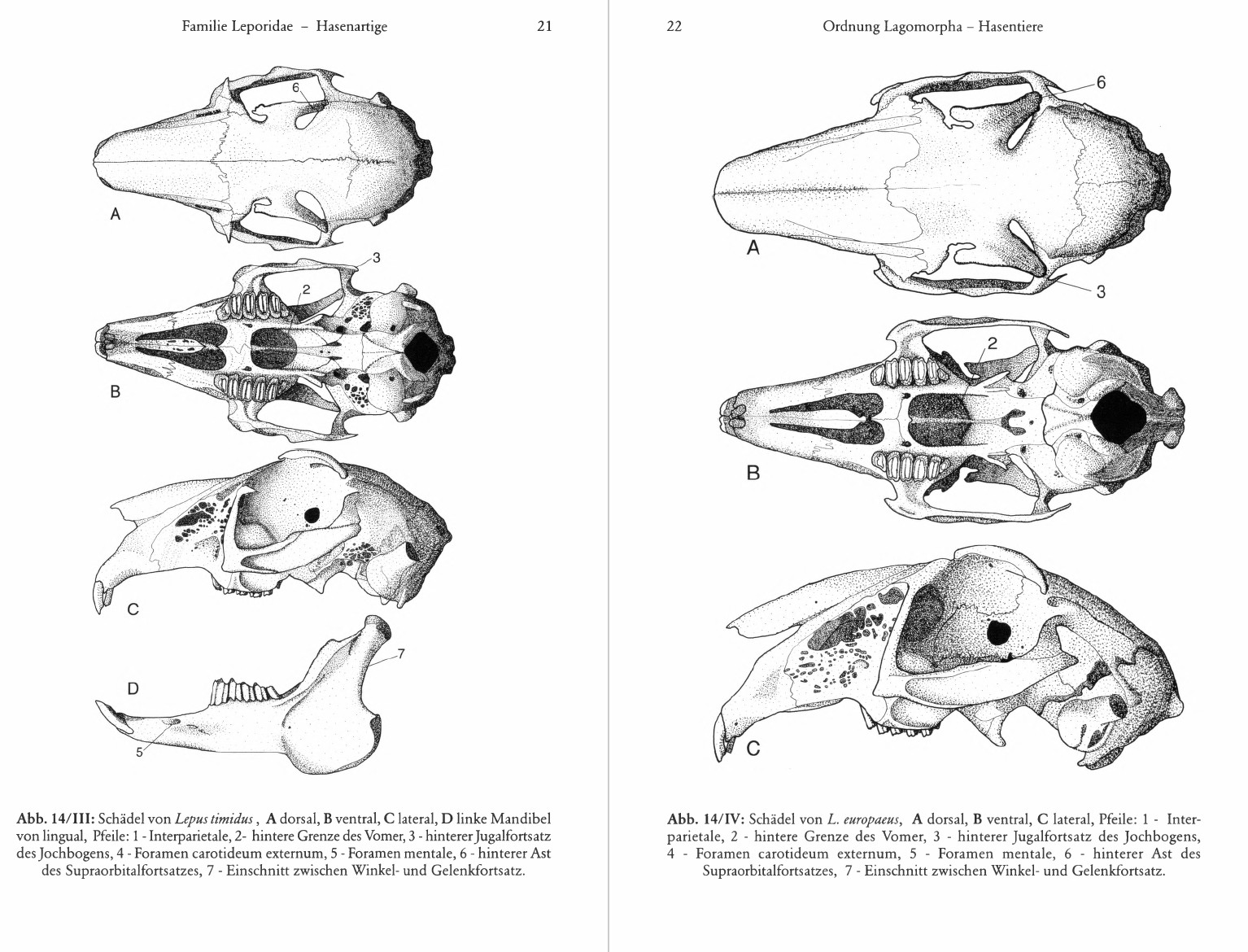
Schädel: Auffällig sind die siebartigen Durchbrüche vor allem an den vorderen, seitlichen Flügeln der Maxillaria, in geringerem Umfang aber auch an Squamosum, Occipitale, Parietale und Interparietale. Von den Stirnbeinen ausgehende Supraorbitalfortsätze überdecken die Augenhöhlen, die vor allem von den großen Augen eingenommen werden, wogegen für den kleinen M. temporalis nur wenig Platz bleibt. Trotz großer innerartlicher Variabilität spielt ihre Gestalt auch bei der Unterscheidung der Arten eine Rolle. Dasselbe gilt für die Breite der Foramina incisiva und der Fossa mesopterygoidea im Verhältnis zueinander, zur Länge des knöchernen Gaumens und des Gesamtschädels. Bei ausdauernd laufenden Leporiden ist die Fossa mesopterygoidea anscheinend breiter als bei Kurzstreckenläufern wie dem Kaninchen. Im Zusammenhang damit dehnen sich auch die Vomeres und das Ende der Nasenscheidewand bei Läufern weiter nach hinten aus. Am Unterkiefer fallen der große, plattenförmige Winkelfortsatz und der hohe, den rudimentären Kronenfortsatz überragende Gelenkfortsatz auf. Zwischen M3 und der Gelenkfläche verläuft eine labial vom Kronenfortsatz begrenzte Rinne. Gehörkapsel und äußerer Gehörgang werden ausschließlich vom Ectotympanicum gebildet.
Quelle: Handbuch der Säugetiere Europas: Hasentiere




